
By Dr Sean Devenish, Chief Scientist, Fluidic Sciences Limited
Protein interactions are central to biology and their characterization is important for scientists from basic research to drug development. Traditionally, protein interaction assays, such as the dominant Surface Plasmon Resonance (SPR) technology, rely on immobilizing proteins on a surface. However, this approach has several limitations. In this article, we will explore these limitations and demonstrate how in-solution approaches can effectively address them.
Why do we study protein interactions?
Proteins are the actors of biology, realizing the potential encoded in the genes of DNA. To coordinate their many and diverse roles, the majority of proteins interact with other proteins. Interaction may be persistent or transient, spontaneous or triggered by stimuli such as post-translational modification. Interaction can result in the formation of simple homomeric species or large, multi-component cellular machinery. It is immediately obvious that protein interactions are critical to biology – without protein interactions life would not exist.
Given the diversity of proteins themselves, their interactions can be challenging to study and understand. There are, nonetheless, compelling reasons to invest the time and effort to explore how proteins interact with each other. Understanding interactions is critical for basic biological research, synthetic biology, studies of health and disease, and in drug development. In basic research, examining similar interactions in different organisms and uncovering networks can improve our understanding of how life operates at a fundamental level. Synthetic biology will benefit from deeper knowledge of complex formation and function switching that commonly centres around protein interactions. Medical research depends heavily on understanding protein interactions as interactions are directly involved in diseases as diverse as cancer and neurodegeneration, autoimmune and infectious diseases. Finally, modern pharmaceutical development is heavily focused on proteins as therapeutics, so developing the full picture of how these drugs interact with their targets is crucial to the development and approval of new therapies.
It is worth taking a moment to consider the parameters that govern protein interactions and that can be determined by experimental methods to shed light on protein social behavior. The first is the precise identity of interacting species. As proteins are subject to a range of post-translational modifications (PTMs), their interactions may be dependent on the presence of particular modifications. In cases where PTMs may play a role, their presence and type should be examined using identity proteomic approaches such as mass spectrometry. Once the species involved in an interaction are known we can start asking questions about the nature of their interaction; how tightly they interact, their interaction affinity, is a key factor as this determines the amount of complex formed given the concentrations of the component species. The kinetics of complexation, or the rate at which complex forms (or disassembles), can be important in some cases where concentrations of interacting species vary over time.
Focusing on the key parameter of interaction affinity, there are a wide range of protein interaction assays that can be used to measure binding strength. These assays can be broadly divided into two groups depending on whether the interaction is measured at a synthetic surface or in solution.
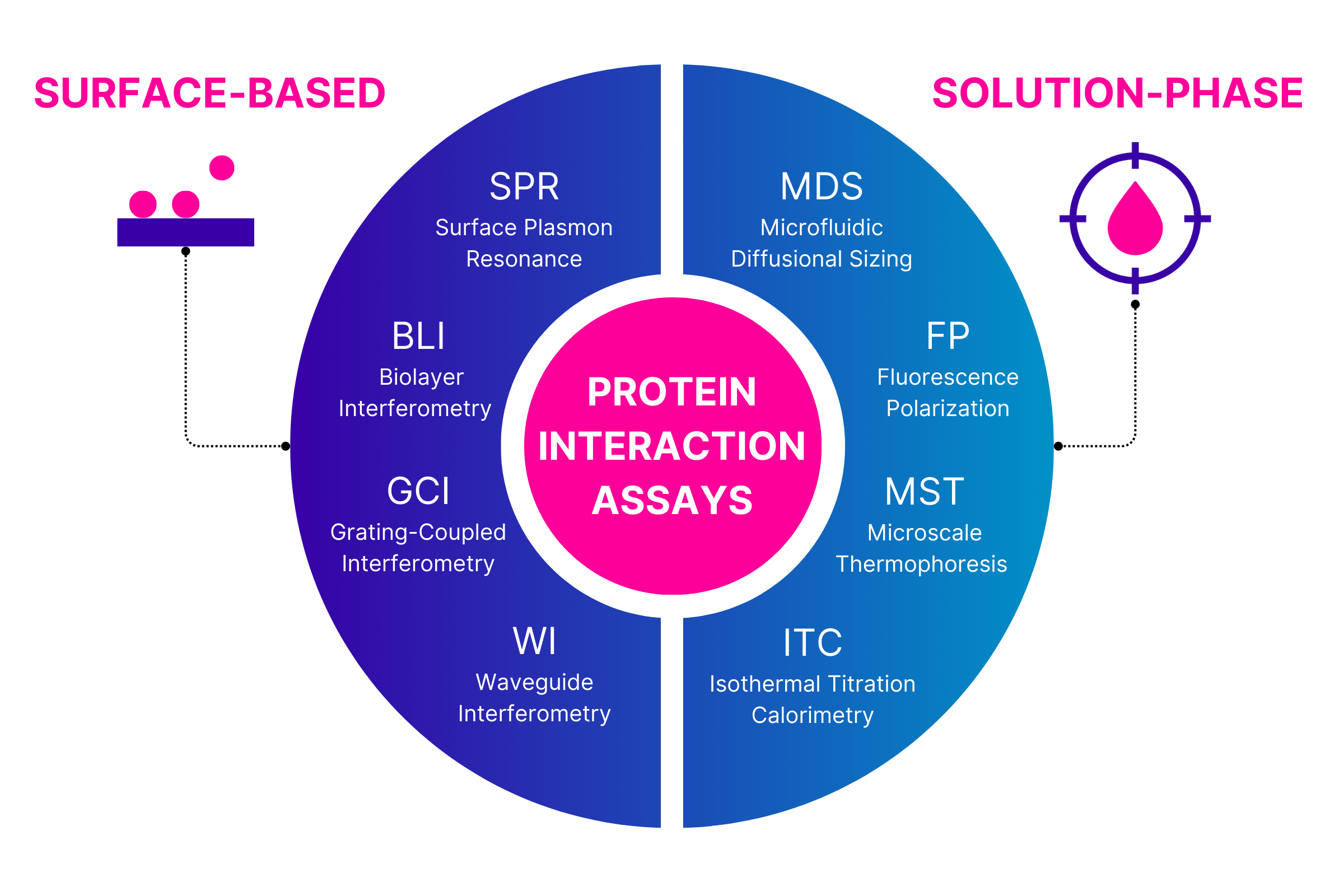
Why the frustration with surface-based protein interaction assays
There are a range of technologies that characterize interactions by detecting interaction at or near a sensor surface. Surface Plasmon Resonance (SPR) is the dominant approach, with several companies offering SPR instruments, but there are a growing number of alternative technologies that operate in a similar fashion although with differences in the detail, including Biolayer Interferometry (BLI), Grating-Coupled Interferometry (GCI), Waveguide Interferometry (WI).
While these technologies all have different ways of detecting a binding signal, they share a common experimental approach. In all cases, a probe of interest - the target - is tethered to a surface. Capture can be covalent or non-covalent by using surfaces pre-functionalized with a tag-specific binder such as streptavidin, protein G, or NTA. The surface is then treated to reduce non-specific binding events and the analyte applied to the surface to allow binding. Once binding is complete the analyte is replaced with analyte-free buffer and dissociation is monitored. Binding and unbinding of the analyte are detected in real time so kinetics are determined. Typically, affinity is calculated from the kinetic on- and off-rates.
Though used regularly, surface-based approaches come with challenges:
Immobilization can be challenging in some cases and may require optimization prior to beginning analytical measurements
Surface attachment can create a range of artefacts and so experiments need to include suitable controls and/or modifications to compensate:
Non-specific binding to the surface is typically a challenge with complex samples such as biofluids but analytes can also bind the surface directly
If target densities are sufficiently high then avid binding can occur for multivalent analytes, artificially increasing the apparent affinity of interaction
Conformational deformation and/or steric occlusion of a target can lead to epitope divergence and multiphasic binding behavior
Solution-phase protein interaction assays: a new approach to overcome limitations in surface-based techniques
Solution-phase analyses offer more biologically relevant measurements as both binding species are free to diffuse in three dimensions, and in some cases are also able to operate directly in biological samples. The key discriminator regarding matrix compatibility is whether the target is labeled or not. Label-free techniques such as Isothermal Titration Calorimetry (ITC) offer the ability to characterize interactions of entirely unmodified proteins, but at the price of operating only in simple buffer matrices. Addition of a fluorescent label to the target is a straightforward and minimal change that provides selectivity and specificity of detection so enables measurements in complex matrices such as culture medium, cell lysate, or even directly in biofluids such as serum or plasma. Examples of technologies able to exploit fluorescent labeled targets include Fluorescence Polarization (FP), Microscale Thermophoresis (MST) and Microfluidic Diffusional Sizing (MDS). So, what are the advantages of in-solution analysis of protein interactions?
Without the need to manage immobilization, assay development is typically much easier for in-solution measurements than surface-based approaches
Working in solution opens the door to analyzing interactions of challenging targets such as intrinsically disordered proteins, protein complexes, and membrane proteins
Biology takes place in solution; solution-phase measurements are more biologically representative than surface-based measurements
Operating in solution means the experimenter has control over the concentrations of both species in a reaction which opens up exciting new analytical options as seen in the Fluidity Insight platform of Fluidic Sciences
Measuring directly in complex matrices such as serum or plasma is possible as non-specific interactions with surfaces are inconsequential when detection takes place in solution
Microfluidic Diffusional Sizing (MDS) can be considered in more detail as an example technology for in-solution interaction assays. MDS, incorporated in the Fluidity One-M instrument by Fluidic Sciences, works on an intuitive and straightforward principle: changes in molecular size are inherent to every protein interaction. MDS measures molecular size (hydrodynamic radius) of a sample by evaluating its diffusion in a microfluidic chamber. The method therefore doesn’t require immobilization or purification, making it possible to work with a wide variety of targets. Among a wide range of studies, MDS has been used to study the interaction of the anti-Alzheimer’s disease therapeutic aducanumab with the challenging targets monomeric and fibrillar Aβ peptide [1], the displacement of α-synuclein oligomers from lipid membranes by monomeric α-synuclein [2], and the dynamic behavior of myosin within cardiac muscle myosin filaments [3].
Going without immobilization and purification also means that you can use MDS to measure the interactions in their native environment, in crude samples, directly in serum or plasma. For example, MDS has been exploited to reveal the impact of immunomodulatory therapy on immune responses to SARS-CoV-2 vaccination [4], characterize secretory antibodies directly in saliva samples [5], and examine affinity of serum alloantibody responses to human leukocyte antigen to better predict and monitor organ transplant outcomes [6].
MDS can also provide measurements of molecular size, stoichiometry and concentration alongside affinity, giving a full picture of binding events. Illustrating the ability of MDS to go beyond classical interaction studies, it has been used to identify small molecule screening hits that induce protein aggregation in a primary assay rather than site-specifically binding the target [7], monitor early stages of phase separation of nucleic acids induced by antimicrobial peptides [8], and study the micelle-forming behavior of the human chaperone protein DNAJB6b [9].
Recommendation
Surface-based and solution-phase technologies for characterizing protein interactions are fundamentally distinct, and so while offering different advantages and disadvantages, operate together to cover a wide range of experimental needs. For any project, it is important to have orthogonal approaches to validate initial results and thereby avoid reporting or making decisions on artefactual data. Thus, surface-based and solution-phase methods for characterizing protein interactions are highly complementary, and would ideally both be employed as standard practice when carrying out the important research into the social behavior of proteins.
About the Author:
Dr Sean Devenish has a PhD in chemistry from the University of Canterbury, New Zealand, and over a decade of postdoctoral experience in biochemistry, biophysics and protein engineering, including six years at the University of Cambridge. Sean has published more than 40 peer-reviewed scientific publications including two in Nature journals. He has been working with Microfluidic Diffusional Sizing for over nine years, contributing to development and optimization of the technology, analytics and assays.
References
Linse, S.; Scheidt, T.; Bernfur, K.; Vendruscolo, M.; Dobson, C.M.; Cohen, S.I.A.; Sileikis, E.; Lundqvist, M.; Qian, F.; O’Malley, T.; et al. Kinetic Fingerprints Differentiate the Mechanisms of Action of Anti-Aβ Antibodies. Nat. Struct. Mol. Biol.2020, 27, 1125–1133. https://doi.org/10.1038/s41594-020-0505-6
2. Šneiderienė, G.; Czekalska, M.A.; Xu, C.K.; Jayaram, A.K.; Krainer, G.; Arter, W.E.; Peter, Q.A.E.; Castellana-Cruz, M.; Saar, K.L.; Levin, A.; et al. α-Synuclein Oligomers Displace Monomeric α-Synuclein from Lipid Membranes. ACS Nano2024, 18, 17469–17482. https://doi.org/10.1021/acsnano.3c10889
3. Kelly, C.M.; Martin, J.L.; Previs, M.J. Myosin Folding Boosts Solubility in Cardiac Muscle Sarcomeres. JCI Insight2024, 9. https://doi.org/10.1172/jci.insight.178131
4. Priddey, A.; Chen-Xu, M.X.H.; Cooper, D.J.; MacMillan, S.; Meisl, G.; Xu, C.K.; Hosmillo, M.; Goodfellow, I.G.; Kollyfas, R.; Doffinger, R.; et al. Microfluidic Antibody Profiling after Repeated SARS-CoV-2 Vaccination Links Antibody Affinity and Concentration to Impaired Immunity and Variant Escape in Patients on Anti-CD20 Therapy. Front. Immunol.2024, 14. https://doi.org/10.3389/fimmu.2023.1296148
5. O’Mahoney, C.; Watt, I.; Fiedler, S.; Devenish, S.; Srikanth, S.; Justice, E.; Dover, T.; Dean, D.; Peng, C. Microfluidic Diffusional Sizing (MDS) Measurements of Secretory Neutralizing Antibody Affinity Against SARS-CoV-2. Ann. Biomed. Eng.2024, 52, 1653–1664. https://doi.org/10.1007/s10439-024-03478-0
6. Schneider, M.M.; Scheidt, T.; Priddey, A.J.; Xu, C.K.; Hu, M.; Meisl, G.; Devenish, S.R.A.; Dobson, C.M.; Kosmoliaptsis, V.; Knowles, T.P.J. Microfluidic Antibody Affinity Profiling of Alloantibody-HLA Interactions in Human Serum. Biosens. Bioelectron.2023, 228, 115196. https://doi.org/10.1016/j.bios.2023.115196
7. Heuser, A.; Abdul Rahman, W.; Bechter, E.; Blank, J.; Buhr, S.; Erdmann, D.; Fontana, P.; Mermet-Meillon, F.; Meyerhofer, M.; Strang, R.; et al. Challenges for the Discovery of Non-Covalent WRN Helicase Inhibitors. ChemMedChem2024, 19, e202300613. https://doi.org/10.1002/cmdc.202300613
8. Sneideris, T.; Erkamp, N.A.; Ausserwöger, H.; Saar, K.L.; Welsh, T.J.; Qian, D.; Katsuya-Gaviria, K.; Johncock, M.L.L.Y.; Krainer, G.; Borodavka, A.; et al. Targeting Nucleic Acid Phase Transitions as a Mechanism of Action for Antimicrobial Peptides. Nat. Commun.2023, 14, 7170. https://doi.org/10.1038/s41467-023-42374-4
9. Carlsson, A.; Olsson, U.; Linse, S. On the Micelle Formation of DNAJB6b. QRB Discov.2023, 4, e6. https://doi.org/10.1017/qrd.2023.4